|
|
Home
People About the Lab Oomycetes Genomics Effectors Publications Web supplements Useful links |
This is the old homepage of Sophien Kamoun's lab at TSL. For the new homepage, please visit http://KamounLab.net. Effectors: Key to understanding pathogenicity
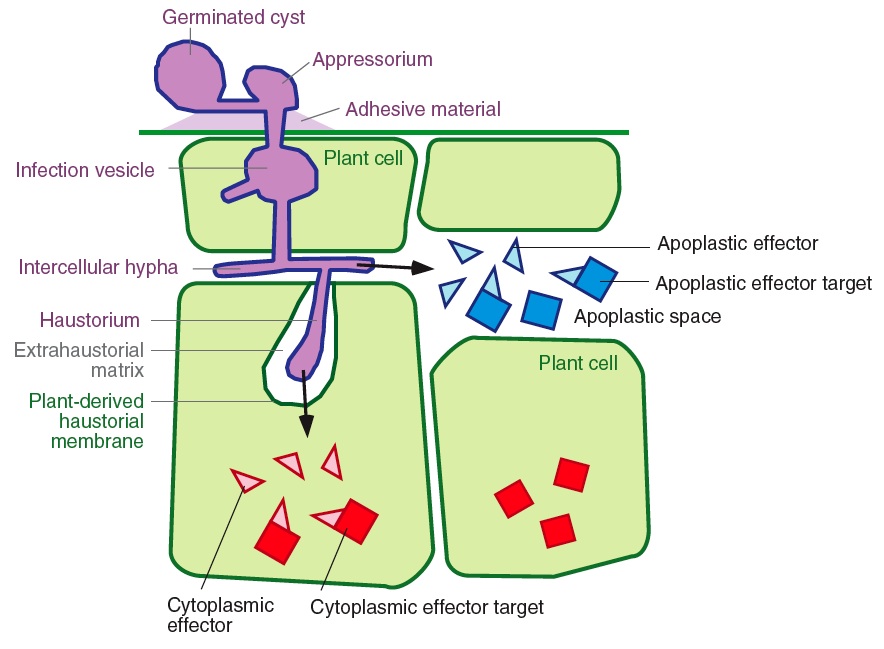
Overview Plant pathogenic bacteria, oomycetes, fungi, and nematodes secrete an arsenal of effector proteins that modulate plant innate immunity and enable parasitic infection. Deciphering the biochemical activities of effectors to understand how pathogens successfully colonize and reproduce on their host plants has become a driving paradigm in the field of molecular plant pathology. In the last three years, unparalleled progress has been made in understanding effector biology in the oomycetes. The identification of a vast repertoire of secreted oomycete proteins and their subsequent characterization led to the classification of effectors based on their target site in the host. Apoplastic effectors are secreted into the plant extracellular space, while cytoplasmic effectors are translocated inside the plant cell, where they target different subcellular compartments. Understanding the perturbations caused by oomycete effectors should prove critical for unravelling mechanisms of pathogenicity, as well as further illuminate novel aspects of plant defense and innate immunity. Of particular interest are the activities of the RXLR and CRN 'Crinkler' families of cytoplasmic effectors that are characterized by conserved motifs following the signal peptide.
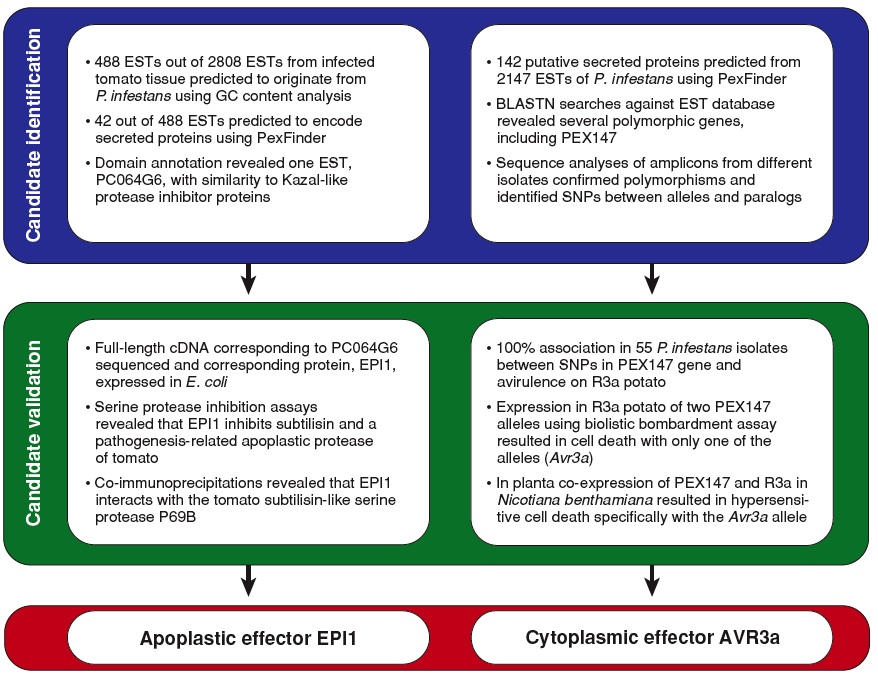
Functional connections between pathogen effectors and host defenses We used data mining of genome sequences and functional genomics to identify several hundred genes that are likely to encode secreted effectors of both classes (see examples of functional genomics pipelines below). Apoplastic effectors are often small cysteine-rich proteins, such as several serine and cysteine protease inhibitors that target host proteases and the phytotoxin-like SCR74. In contrast, cytoplasmic effectors belong to the RXLR and CRN families that comprises over 200 members each in P. infestans. One RXLR effector, AVR3a, triggers the hypersensitive response (HR) in R3a plants and suppresses the HR induced by INF1 elicitin. Our immediate goals are to understand the molecular function and biochemical activity of apoplastic and cytoplasmic effectors of P. infestans and other oomycetes:
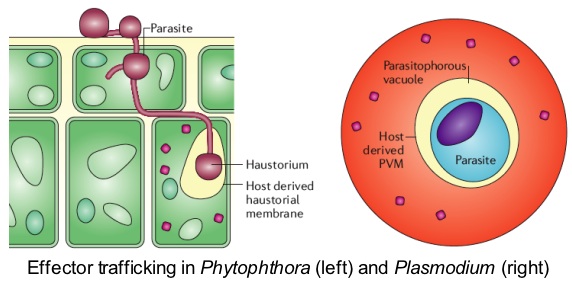
Effector protein trafficking in eukaryotic pathogens There is emerging evidence that delivery of oomycete proteins inside plant cells may take place, possibly through infection vesicles and haustoria. For example, a common feature of all known oomycete Avr proteins, such as AVR3a, is that they function inside host cells. We study the mechanisms of effector protein translocation inside host cells, arguably one of the most critical questions in the study of eukaryotic plant pathogens. Recent findings offer new avenues on how to tackle this problem. RXLR cytoplasmic effectors are characterized by a highly conserved N-terminal sequence motif that is positioned within sixty amino acids of the signal sequence. This motif appears to function in translocation of effectors into the host cytoplasm. Indeed, these Phytophthora motifs are similar to and functionally interchangeable with a host-targeting signal that is required for translocation of proteins from malaria parasites (Plasmodium species) into the cytoplasm of host cells. These findings raise the possibility that plant and animal eukaryotic pathogens share similar mechanisms for effector secretion inside host cells. We are exploiting these findings to study the machinery of effector trafficking using structure-function and cellular localization analyses.
Review articles Morgan, W., and Kamoun, S. 2007. RXLR effectors of plant pathogenic oomycetes. Current Opinion in Microbiology, in press. Kamoun, S. 2007. Groovy times: filamentous pathogen effectors revealed. Current Opinion in Plant Biology, 10:358-365. Supplementary Material. iTunes iMix companion playlist. Haldar, K., Kamoun, S., Hiller, L. N., Bhattacharjee, S., and and van Ooij, C. 2006. Common infection strategies of pathogenic eukaryotes. Nature Reviews Microbiology, 4:922-931. Kamoun, S. 2006. A catalogue of the effector secretome of plant pathogenic oomycetes. Annual Review of Phytopathology, 44: 41-60. Birch, P. R. J., Rehmany, A. P., Pritchard, L., Kamoun, S., and Beynon, J. L. 2006. Trafficking arms: oomycete effectors enter host plant cells. Trends in Microbiology, 14:8-11. Other relevant articles Win, J, Morgan, W., Bos, J., Krasileva, K.V., Cano, L.M., Chaparro-Garcia, A., Ammar, R., Staskawicz, B.J., and Kamoun, S. 2007. Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell, 19:2349-2369. Supplementary Material. Bos, J..I.B., Kanneganti, T. -D., Young, C., Cakir, C., Huitema, E., Win, J., Armstrong, M., Birch, P.R.J., and Kamoun, S. 2006. The C-terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in Nicotiana benthamiana. Plant Journal, 48:165-176. Supplementary Material. Bhattacharjee, S., Hiller, L. N., Liolios, K., Win, J., Kanneganti, T. -D., Young, C., Kamoun, S., and Haldar, K. 2006. The malarial host-targeting signal is conserved in the Irish potato famine pathogen. PLoS Pathogens 2:e50. Tian, M., Benedetti, B., and Kamoun, S. 2005. A second Kazal-like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis-related protease P69B of tomato. Plant Physiology, 138:1785-93. Armstrong, M.R., Whisson, S.C., Pritchard, L., Bos, J.I.B., Venter, E., Avrova, A.O., Rehmany, A.P., Bšhme, U., Brooks, K., Cherevach, I., Hamlin, N., White, B., Fraser, A., Lord, A., Quail, M.A., Churcher, C., Hall, N., Berriman, M., Huang, S., Kamoun, S., Beynon, J.L., and Birch, P.R.J. 2005. An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proceedings of the National Academy of Sciences USA, 102:7766-7771. Rehmany, A. P., Gordon, A., Rose, L.E., Allen, R.L., Armstrong, M.R., Whisson, S.C., Kamoun, S., Tyler, B.M., Birch, P.R.J., and Beynon, J.L. 2005. Differential recognition of highly divergent downy mildew avirulence gene alleles by RPP1 genes from two Arabidopsis lines. Plant Cell, 17:1839-1850. Liu, Z., Bos, J.I.B., Armstrong, M., Whisson, S.C., da Cunha, L., Torto-Alalibo, T., Win, J., Avrova, A.O., Wright, F., Birch P.R.J., and Kamoun, S. 2005. Patterns of diversifying selection in the phytotoxin-like scr74 gene family of Phytophthora infestans. Molecular Biology and Evolution, 22:659-672. Supplementary Material. GenBank accessions. Tian, M., Huitema, E., da Cunha, L., Torto-Alalibo, T., and Kamoun, S. 2004. A Kazal-like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease P69B. Journal of Biological Chemistry, 279:26370-26377. GenBank accessions. |